健康好帮手

内容仅限于知识科普,不代表对本公司产品的宣传。
在学术上研究比较多的氢化酶是细菌氢化酶,研究氢化酶的一个主要理由是希望通过仿生学建立生物制造氢气的技术,例如模仿光合细菌合成氢气,这已经成为氢气能源领域的一个重要研究领域。这一领域的研究,也给我们了解氢化酶的相关信息提供了重要基础。因此,要研究植物氢化酶或理解动物细胞合成利用氢气的可能分子基础,都应该认真学习和掌握细菌氢化酶相关知识。或者从事氢气生物医学研究的学者,都应该了解细菌氢气代谢的相关知识。这类根据最新综述内容,结合我自己的理解,对细菌氢化酶相关研究进行讨论。
我过去曾经对细菌氢化酶的有关知识进行过介绍,也强调或直接提出过细菌氢化酶和氢气医学研究相关性的观点。1、线粒体整体进化目前流行观点是氢假说,就是线粒体来源就是一种产氢气微生物,而宿主菌则是利用氢气的厌氧菌。线粒体复合物I在进化上来源于细菌氢化酶,理论上具有合成氢气的潜力,更大可能是氢气发挥调节作用的分子目标。2、细胞内存在多种氢化酶同源分子,例如铁硫簇相关蛋白,这些蛋白都具有电子传递的特征,符合和氢气代谢的相关性逻辑,其活性也可能受到氢气的调节。3、真核细胞本身可能具有合成氢气的潜力,如果能证实,则能极大提高氢气的生物学地位。(氢思语阅读最新文献的感言。)4、应该利用目前了解的氢化酶基因序列,全面分析人体内相关基因,并对这些基因和氢气代谢的相关性进行研究,有希望发现这些蛋白可能受氢气调节,甚至有可能这些基因仍然保留代谢氢气的潜力。
绿藻是一种真核生物,含有叶绿体具有光合作用功能,经常被作为高等植物生理生化功能研究的模式生物。莱茵衣藻(Chlamydomonas reinhardtii)是一种单细胞真核绿藻,其基因组测序已全部完成,并成功实现了细胞核、叶绿体、线粒体基因组的遗传转化。莱茵衣藻有两个鞭毛和一个占据细胞内大部分空间的单个叶绿体。莱茵衣藻是一种非常有价值的真核模式生物,被广泛用于与光合作用、呼吸作用、脂类合成、细胞运动(生物鞭毛)、非生物胁迫等生物学过程相关的功能研究。
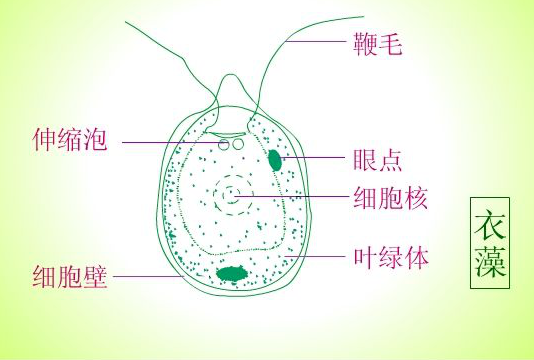
莱茵衣藻的结构,一个大叶绿体两个鞭毛。
衣藻不是高等植物,但这种生物具有制造氢气的能力,莱茵衣藻有两种铁铁氢化酶HYDA1和 HYDA2,大小分别拥有497和505个氨基酸残基。有意思的是,几乎所有研究表明,藻类都拥有这两类氢化酶。氢化酶位于基质能和光合作用电子传递偶合,从铁氧还蛋白接受电子。光合系统II可以提供电子,但是产生氧气会抑制氢化酶活性。这种活性被NAD(P)H-质体醌氧化还原酶驱动。氢化酶基因在无氧条件下表达增强。莱茵衣藻氢气产量还依赖于光线,在无二氧化碳和低硫情况下氢气产量增加。根据这些现象推测氢气产生依赖于光合系统II,产生淀粉的后续产物,是电子受体的产物。但是缺乏二磷酸核酮糖羧化酶时产生淀粉障碍,但这类细胞仍然可能通过光系统II产生氢气。Torzillo等发现,硫剥夺莱茵衣藻活体氢气产生速率为10nmol/h.µg叶绿素II。光系统II中心蛋白D1突变可以增加氢气产生和持续产生时间。Philipps等研究发现,氮源缺乏和硫源缺乏时,虽然淀粉积聚不受影响,硫源缺乏导致的氢气产量增加会下降。提示因为细胞色素b6f复合物降解增加,导致铁硫蛋白水平下降。
低等光合生物如莱茵衣藻在某些特定环境条件下产生大量氢气,产生的氢气生理作用可能是帮助在氧气浓度升高时协调电子流稳定。是避免这些电子被氧气分子抢夺?产生的氢气容易扩散,相当于电子的泄露,持续产生氢气显然有浪费能量的嫌疑。但是对于人类来说,充分利用这种天然的光合产氢气能力,可以源源不断制造绿色氢气能源。
使用 HYDA1氨基酸序列在BLAST上检索显示,拥有氢化酶蛋白同源序列的衣藻有许多,包括简单四豆藻、 胸状团藻、盘藻、小球藻等。这说明衣藻产生是普遍现象,不只是莱茵衣藻特有。
如果氢气活性氧和一氧化氮代谢有影响,那么对氢化酶的调节可能有更深层意义,理论上活性氧和一氧化氮对氢化酶活性有反馈调节效应。确实早有研究报道,一氧化氮能抑制氢化酶活性氧。
一氧化氮能对蛋白质进行S-亚硝基化将半胱氨酸巯基修饰成S-亚硝酰基,或者将酪氨酸修饰成硝基酪氨酸。氢化酶HYDA1有11个,HYDA2有9个半胱氨酸残基,其中6个同源。如果这些巯基没有深埋在蛋白三维结构内部,都有可能被一氧化氮或活性氧修饰。利用iSNO-PseAAC预测工具分析发现,HYDA1的Cys170、Cys191和Cys225可发生S亚硝基化修饰,HYDA2的Cys17、Cys33、Cys194和 Cys228可发生S亚硝基化修饰。两类氢化酶共同可被修饰的残基有两个,Cys191/194和Cys225/228,提示这非常有可能是一氧化氮调节部位。但是围绕在亚硝基化半胱氨酸周围的结构比一级结构序列更重要,研究提示(IL)-X-C-X-X-(DE)序列是形成亚硝基化修饰的关键结构,两种氢化酶都缺乏这个序列,提示它们并不能被亚硝基化调节。当然对氢化酶活性的调节不一定发生在氨基酸残基层次。早就有研究发现一氧化氮对氢化酶活性有抑制作用,最近对脱磷孤菌属氢化酶的这一现象进行了研究。
Ceccaldi等发现氢化酶金属元素是一氧化氮的作用目标,一氧化氮能抑制氢化酶催化活性中心,也能不可逆破坏铁硫簇中心,这可能类似一氧化碳和血红素铁共价结合阻断运输氧气的情况。从目前了解的资料看,一氧化氮对巯基修饰并不具有任何选择性,活性氧对巯基的修饰尚无研究报道。所以,氢化酶受活性氧一氧化氮调节方面仍然需要进一步探讨。同样,对硫化氢调节目标蛋白的模式也缺乏了解,根据硫化氢效应基础和亚硝基化类似的情况推测,氢化酶受硫化氢调节目标也可能是巯基氨基酸和酪氨酸。
Russell G, Zulfiqar F, Hancock JT.Hydrogenases and the Role of Molecular Hydrogen in Plants.Plants (Basel). 2020 Sep 2;9(9):E1136.
以上内容摘自《孙学军 氢思语》,仅限于知识科普,不代表对本公司产品的宣传。








